
Tartalom
- Genetikai információk a prokariótákban
- A gén expresszió szakaszai
- Transzkripció baktériumsejtekben
- Átírás: Beindítási szakasz
- Átírás: meghosszabbítási szakasz
- Átírás: Megszakítási szakasz
- Fordítás baktériumsejtekben
- Fordítás: Bevezetés
- Fordítás: Nyúlás
- Fordítás: Felmondás
- Fordítás és antibiotikumok
- Transzláció utáni fehérjefeldolgozás
- Fehérje-foszforiláció
- Fehérjecetilezés és glikozilezés
- Gén kifejezés Archaea-ban
A prokarióták kicsi, egysejtű élő szervezetek. Ezek a két általános sejttípus egyikét képezik: prokarióta és eukarióta.
Mivel a prokarióta sejtekben nincs sejtmag vagy sejtek, a génexpresszió a nyílt citoplazmában fordul elő, és az összes stádium egyszerre fordulhat elő. Noha a prokarióták egyszerűbbek, mint az eukarióták, a gén expressziójának ellenőrzése továbbra is döntő jelentőségű sejt viselkedésük szempontjából.
Genetikai információk a prokariótákban
A prokarióták két területe a baktériumok és az Archaea. Mindkettőnek nincs meghatározott magja, de genetikai kódjuk és nukleinsavaik vannak. Noha nincsenek olyan komplex kromoszómák, mint amilyeneket eukarióta sejtekben látnánk, a prokarióták kör alakú dezoxiribonukleinsav (DNS) darabokat tartalmaznak a nukleoidban.
A genetikai anyag körül azonban nincs membrán. Általában a prokarióták kevesebb nem kódoló szekvenciát mutatnak a DNS-ben, mint az eukarióták. Ennek oka lehet, hogy a prokarióta sejtek kisebbek és kevesebb helyet foglalnak el a DNS-molekulához.
A nukleoid egyszerűen az a régió, ahol a DNS él a prokarióta sejtben. Szabálytalan alakú és méretétől függően változhat. Ezenkívül a nukleoid kapcsolódik a sejtmembránhoz.
A prokarióták körkörös DNS-sel is rendelkezhetnek plazmidok. Lehetséges, hogy egy vagy több plazmidot tartalmaznak egy sejtben. A sejtosztódás során a prokarióták átjuthatnak a DNS-szintézisbe és a plazmidok elválasztásába.
Az eukarióta kromoszómáival összehasonlítva a plazmidok általában kisebbek és kevesebb DNS-sel rendelkeznek. Ezenkívül a plazmidok önmagukban is replikálódhatnak más celluláris DNS nélkül. Egyes plazmidok nem esszenciális gének kódját hordozzák, például azok, amelyek baktériumok antibiotikum-rezisztenciáját adják.
Bizonyos esetekben a plazmidok képesek mozogni az egyik sejtből a másikba és olyan információkat megosztani, mint például az antibiotikum-rezisztencia.
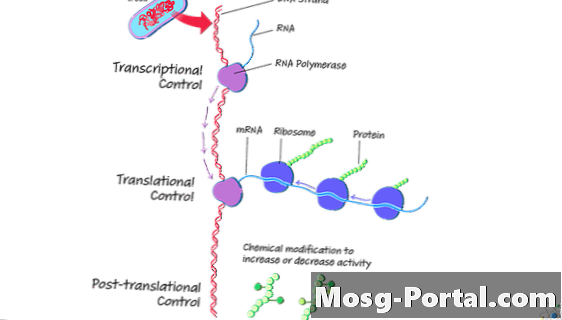
A gén expresszió szakaszai
A gén expresszió az a folyamat, amelyen keresztül a sejt a genetikai kódot aminosavakká alakítja át fehérjetermelés céljából. Az eukariótákkal ellentétben a két fő szakasz, az átírás és a fordítás, prokariótákban egyszerre történhet.
A transzkripció során a sejt transzlálja a DNS-t Messenger RNS (mRNS) molekulává. A transzláció során a sejt az aminosavakat állítja elő az mRNS-ből. Az aminosavak alkotják a fehérjéket.
Mind az átírás, mind a fordítás a prokariótákban történik citoplazma. Ha mindkét folyamat egyszerre zajlik le, a sejt nagy mennyiségű fehérjét termelhet ugyanabból a DNS-templátból. Ha a sejtnek már nincs szüksége a fehérjére, akkor a transzkripció leállhat.
Transzkripció baktériumsejtekben
A transzkripció célja egy komplementer ribonukleinsav (RNS) szál létrehozása egy DNS-templátból. A folyamat három részből áll: iniciálás, lánc meghosszabbítás és befejezés.
Annak érdekében, hogy a beindítási szakasz megtörténjen, a DNS-nek először le kell lazulnia, és a terület, ahol ez megtörténik, a transzkripciós buborék.
A baktériumokban ugyanazt az RNS-polimerázt találja, amely felelős az összes transzkripcióért. Ennek az enzimnek négy alegysége van. Az eukariótákkal ellentétben a prokarióták nem rendelkeznek transzkripciós faktorokkal.
Átírás: Beindítási szakasz
A transzkripció akkor kezdődik, amikor a DNS lazul, és az RNS polimeráz kötődik egy promoterhez. A promoter egy speciális DNS-szekvencia, amely egy adott gén elején létezik.
Baktériumokban a promoternek két szekvenciája van: -10 és -35 elem. A -10 elem az a pont, ahol a DNS általában kilazul, és 10 nukleotid távolságra van az iniciációs helytől. A -35 elem 35 nukleotid a helytől.
Az RNS-polimeráz az egyik DNS-szálra támaszkodik templátként, mivel egy új RNS-szálat épít, amelyet RNS-transzkripciónak hívnak. A kapott RNS-szál vagy primer transzkriptum majdnem megegyezik a nem templátos vagy kódoló DNS-szálral. Az egyetlen különbség az, hogy az összes timin (T) bázis uracil (U) bázis az RNS-ben.
Átírás: meghosszabbítási szakasz
A transzkripció lánchosszabbítási fázisa során az RNS-polimeráz a DNS-templát szál mentén mozog, és mRNS-molekulát készít. Az RNS szál hosszabbodik, mivel több nukleotidot adunk hozzá.
Ennek megvalósításához lényegében az RNS-polimeráz a DNS-állvány mentén sétál 3–5 irányban. Fontos megjegyezni, hogy a baktériumok képesek létrehozni polycistronic mRNS-ek ez a kód több fehérjére.
••• TudományÁtírás: Megszakítási szakasz
A transzkripció befejező szakaszában a folyamat leáll. A prokariótákban kétféle terminációs fázis létezik: Rho-függő és Rho-független termináció.
Ban ben Rho-függő lezárás, egy speciális Rho nevű protein faktor megszakítja a transzkripciót és megszünteti azt. A Rho protein faktor egy specifikus kötőhelyen kapcsolódik az RNS szálhoz. Ezután a szál mentén mozog, hogy elérje az RNS polimerázt a transzkripciós buborékban.
Ezután Rho leválasztja az új RNS szálat és a DNS templátot, így a transzkripció véget ér. Az RNS polimeráz leáll, mert eléri a kódoló szekvenciát, amely a transzkripció stop pontja.
Ban ben Rho-független lezárás, az RNS molekula hurkot képez és levál. Az RNS-polimeráz a templát-szálon olyan DNS-szekvenciát ér el, amely a terminátor, és sok citozin (C) és guanin (G) nukleotidot tartalmaz. Az új RNS-szál hajcsavarodásra hajlik fel. C és G nukleotidjai kötődnek. Ez a folyamat megakadályozza az RNS-polimeráz mozgását.
Fordítás baktériumsejtekben
A transzláció a transzkripció során létrehozott RNS-templát alapján fehérjemolekulát vagy polipeptidet hoz létre. Baktériumokban a fordítás azonnal megtörténhet, és néha a transzkripció során kezdődik. Ez azért lehetséges, mert a prokarióták nem tartalmaznak sejtmembránt vagy semmilyen organellát a folyamatok elválasztására.
Az eukariótákban a dolgok másképp vannak, mert a transzkripció a magban fordul elő, a transzláció pedig a magban citoszóltvagy a sejt intracelluláris folyadéka. Az eukarióta érett mRNS-t is használ, amelyet transzláció előtt dolgozunk fel.
Egy másik ok, amiért a transzláció és a transzkripció egyidejűleg megtörténhet baktériumokban, az az, hogy az RNS-nek nincs szüksége az eukariótákban megfigyelt különleges feldolgozásra. A baktérium RNS azonnal készen áll a transzlációra.
Az mRNS-szál nukleotidcsoportokat hív kodon. Minden kodonnak három nukleotidja van, és egy adott aminosav-szekvenciát kódolnak. Bár csak 20 aminosav van, a sejtekben 61 aminosav kodon van és három stop kodon található. Az AUG a kezdő kodon, és elkezdi a fordítást. Ezenkívül a metionin aminosavat kódolja.
Fordítás: Bevezetés
A transzláció során az mRNS szál mint templát szolgál aminosavak előállításához, amelyek fehérjévé válnak. A sejt dekódolja az mRNS-t ennek megvalósításához.
Az inicializáláshoz transzfer RNS (tRNS), riboszóma és mRNS szükséges. Minden tRNS-molekula rendelkezik antikodont aminosavra. Az antikodon komplementer a kodonnal. Baktériumokban a folyamat akkor kezdődik, amikor egy kis riboszómális egység kapcsolódik az mRNS-hez a Shine-Dalgarno sorozat.
A Shine-Dalgarno szekvencia egy speciális riboszómális kötési terület mind a baktériumokban, mind az archaában. Általában nyolc nukleotidot látszik az AUG startkodonból.
Mivel a baktériumgének transzkripciója csoportokban történhet, egy mRNS sok gént kódolhat. A Shine-Dalgarno szekvencia megkönnyíti a kezdő kodon megtalálását.
Fordítás: Nyúlás
A megnyúlás során az aminosavak lánca hosszabb lesz. A tRNS-ek aminosavakat adnak a polipeptidlánc létrehozásához. A tRNS megkezdi a működését P site, amely a riboszóma középső része.
A P oldal mellett található a Egy oldal. A kodonnal megegyező tRNS eljuthat az A-helyre. Ezután peptidkötés alakulhat ki az aminosavak között. A riboszóma az mRNS mentén mozog, és az aminosavak láncot képeznek.
Fordítás: Felmondás
A megszűnés a stop kodon miatt történik. Amikor egy stop kodon belép az A helyre, a transzlációs folyamat leáll, mert a stop kodonnak nincs komplementer tRNS-je. Fehérjék hívtak felszabadulási tényezők amelyek a P helyhez illeszkednek, felismerik a stop kodonokat és megakadályozzák a peptidkötések kialakulását.
Ez azért történik, mert a felszabadulási tényezők miatt az enzimek hozzáadhatnak egy vízmolekulát, ami elválasztja a láncot a tRNS-től.
Fordítás és antibiotikumok
Ha antibiotikumokat szed egy fertőzés kezelésére, akkor ezek működhetnek úgy, hogy megzavarják a baktériumok transzlációs folyamatát. Az antibiotikumok célja a baktériumok megölése és a szaporodás megakadályozása.
Ennek egyik módja a baktériumsejtek riboszómáinak befolyásolása. A gyógyszerek befolyásolhatják az mRNS transzlációját, vagy blokkolhatják a sejt képességét peptidkötések létrehozására. Az antibiotikumok kötődhetnek a riboszómákhoz.
Például, egyfajta tetraciklinnek nevezett antibiotikum bejuthat a baktériumsejtbe azáltal, hogy átmegy a plazmamembránon és felépül a citoplazmában. Ezután az antibiotikum kötődik a riboszómához és blokkolja a transzlációt.
Egy másik, ciprofloxacin nevű antibiotikum befolyásolja a baktériumsejtet azáltal, hogy egy olyan enzimet céloz meg, amely felelős a DNS lefejtéséért a replikáció lehetővé tétele érdekében. Mindkét esetben megkímélik az emberi sejteket, ami lehetővé teszi az emberek számára, hogy antibiotikumaikat használják anélkül, hogy megölték volna a saját sejteket.
Kapcsolódó téma: többsejtű organizmusok
Transzláció utáni fehérjefeldolgozás
A transzláció vége után néhány sejt folytatja a fehérjék feldolgozását. Poszt-transzlációs módosítások A fehérjék (PTM-ek) lehetővé teszik a baktériumok számára, hogy alkalmazkodjanak környezetükhöz, és szabályozzák a sejtek viselkedését.
Általában véve a PTM-ek ritkábban fordulnak elő a prokariótákban, mint az eukariótokban, de néhány szervezetben vannak ezek. A baktériumok módosíthatják a fehérjéket és megfordíthatják a folyamatokat is. Ez nagyobb sokoldalúságot biztosít, és lehetővé teszi számukra a fehérje módosítást a szabályozásban.
Fehérje-foszforiláció
Fehérje foszforilációja a baktériumokban gyakori módosítás. Ez a folyamat magában foglalja egy foszfátcsoport hozzáadását a fehérjéhez, amely foszfor- és oxigénatomokat tartalmaz. A foszforiláció elengedhetetlen a fehérje működéséhez.
A foszforilezés azonban átmeneti lehet, mivel visszafordítható. Egyes baktériumok a foszforilezést a folyamat részeként használhatják más szervezetek megfertőzésére.
A szerin, treonin és tirozin aminosav oldalláncán bekövetkező foszforilációt nevezzük Ser / Thr / Tyr foszforilezés.
Fehérjecetilezés és glikozilezés
A foszforilált proteinek mellett baktériumok is lehetnek acetilezett és glikozilezett fehérjéket. Lehetnek metilezéssel, karboxilezéssel és más módosításokkal is. Ezek a módosítások fontos szerepet játszanak a sejtek jelátvitelében, szabályozásában és a baktériumokban zajló egyéb folyamatokban.
Például a Ser / Thr / Tyr foszforiláció segít a baktériumoknak reagálni a környezetük változásaira, és növeli a túlélés esélyét.
A kutatások azt mutatják, hogy a sejtek anyagcsere-változásai a Ser / Thr / Tyr foszforilációval járnak, ami azt jelzi, hogy a baktériumok reagálhatnak környezetükre a sejtfolyamatok megváltoztatásával. Ezenkívül a poszt-transzlációs módosítások segítenek gyorsan és hatékonyan reagálni. A változások visszafordításának képessége szintén jelentős ellenőrzést biztosít.
Gén kifejezés Archaea-ban
Az Archaea gén expressziós mechanizmusokat alkalmaz, amelyek jobban hasonlítanak az eukariótákhoz. Noha az archaea prokarióták, vannak néhány közös vonásuk az eukariótákkal, például a gén expressziója és a génszabályozás. Az archaea transzkripciójának és transzlációjának folyamatainak is vannak hasonlóságai a baktériumokkal.
Például mind az archaea, mind a baktériumok metionin az első aminosav, és AUG mint a kezdő kodon. Másrészt, mind az archaea, mind az eukarióta a TATA mező, amely egy olyan DNS-szekvencia a promóter területén, amely megmutatja, hol kell dekódolni a DNS-t.
Az archaea transzlációja hasonló a baktériumokban tapasztalt folyamathoz. Mindkét típusú organizmusnak riboszómái vannak, amelyek két egységből állnak: a 30S és 50S alegységekből. Ezen felül mindkettőnek policisztronos mRNS-ekkel és Shine-Dalgarno szekvenciákkal rendelkezik.
A baktériumok, az archaea és az eukarióták között számos hasonlóság és különbség van. Ugyanakkor mind a génexpresszióra és a génszabályozásra támaszkodnak a túlélés érdekében.